
Molecular Research on Stress Responses in Quercus spp.: From Classical Biochemistry to Systems Biology through Omics Analysis


Abstract
:1. Introduction
2. Classical Biochemistry
3. DNA-Based Markers
4. Genomics
5. Epigenomics
6. Transcriptomics
7. Proteomics
8. Metabolomics
9. Integration and Systems Biology
10. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Nixon, K. Infrageneric classification of Quercus (Fagaceae) and typification of sectional names. Ann. For. Sci. 1993, 50, 25s–34s. [Google Scholar] [CrossRef] [Green Version]
- Manos, P.; Doyle, J.; Nixon, K. Phylogeny, biogeography, and processes of molecular differentiation in Quercus subgenus Quercus (Fagaceae). Mol. Phylogenet. Evol. 1999, 12, 333–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto-Correia, T.; Azeda, C. Public policies creating tensions in Montado management models: Insights from farmers’ representations. Land Use Policy 2017, 64, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Seale, D. Michael Williams: Deforesting the Earth: From prehistory to global crisis, an abridgment. J. Agric. Environ. Ethics 2011, 24, 673–686. [Google Scholar] [CrossRef]
- FAO; UNEP. The State of the World’s Forests 2020: Forests, Biodiversity and People; FAO: Rome, Italy, 2020. [Google Scholar]
- Grantham, H.S.; Duncan, A.; Evans, T.D.; Jones, K.R.; Beyer, H.L.; Schuster, R.; Walston, J.; Ray, J.C.; Robinson, J.G.; Callow, M.; et al. Anthropogenic modification of forests means only 40% of remaining forests have high ecosystem integrity. Nat. Commun. 2020, 11, 5978. [Google Scholar] [CrossRef]
- Wardlaw, T.; Mitchell, P.; Pinkard, L. Combined stresses in forests. In Combined Stresses in Plants; Mahalingam, R., Ed.; Springer: Cham, Switzerland, 2015; pp. 223–244. ISBN 978-3-319-07898-4. [Google Scholar]
- Denman, S.; Doonan, J.; Ransom-Jones, E.; Broberg, M.; Plummer, S.; Kirk, S.; Scarlett, K.; Griffiths, A.R.; Kaczmarek, M.; Forster, J.; et al. Microbiome and infectivity studies reveal complex polyspecies tree disease in Acute Oak Decline. ISME J. 2018, 12, 386–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, N.; Jeger, M.; Kirk, S.; Xu, X.; Denman, S. Spatial and temporal patterns in symptom expression within eight woodlands affected by Acute Oak Decline. For. Ecol. Manag. 2016, 360, 97–109. [Google Scholar] [CrossRef]
- Brown, N.; Vanguelova, E.; Parnell, S.; Broadmeadow, S.; Denman, S. Predisposition of forests to biotic disturbance: Predicting the distribution of Acute Oak Decline using environmental factors. For. Ecol. Manag. 2018, 407, 145–154. [Google Scholar] [CrossRef]
- Denman, S.; Webber, J. Oak declines: New definitions and new episodes in Britain. Q. J. For. 2009, 103, 285–290. [Google Scholar]
- Denman, S.; Brown, N.; Kirk, S.; Jeger, M.; Webber, J. A description of the symptoms of Acute Oak Decline in Britain and a comparative review on causes of similar disorders on oak in Europe. Forestry 2014, 87, 535–551. [Google Scholar] [CrossRef] [Green Version]
- Brady, C.; Arnold, D.; McDonald, J.; Denman, S. Taxonomy and identification of bacteria associated with acute oak decline. World J. Microbiol. Biotechnol. 2017, 33, 143. [Google Scholar] [CrossRef]
- Brown, N.; Jeger, M.; Kirk, S.; Williams, D.; Xu, X.; Pautasso, M.; Denman, S. Acute Oak Decline and Agrilus biguttatus: The Co-occurrence of stem bleeding and D-shaped emergence holes in Great Britain. Forests 2017, 8, 87. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C. Phytophthora cinnamomi and oak decline in southern Europe. Environmental constraints including climate change. Ann. For. Sci. 1996, 53, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Gentilesca, T.; Camarero, J.; Colangelo, M.; Nolè, A.; Ripullone, F. Drought-induced oak decline in the western Mediterranean region: An overview on current evidences, mechanisms and management options to improve forest resilience. iForest Biogeosci. For. 2017, 10, 796–806. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Gómez, F.; Pérez-de-Luque, A.; Sánchez-Cuesta, R.; Quero, J.; Cerrillo, R.M.N. Differences in the response to acute drought and Phytophthora cinnamomi rands infection in Quercus ilex L. seedlings. Forests 2018, 9, 634. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Gómez, F.; Pérez-de-Luque, A.; Navarro-Cerrillo, R. The involvement of Phytophthora root rot and drought stress in Holm oak Decline: From ecophysiology to microbiome influence. Curr. For. Rep. 2019, 5, 251–266. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Natalini, F.; Cardillo, E. Holm oak death is accelerated but not sudden and expresses drought legacies. Sci. Total Environ. 2021, 754, 141793. [Google Scholar] [CrossRef]
- Luque, J.; Parladé, J.; Péra, J. Pathogenicity of fungi isolated from Quercus suber in Catalonia (NE Spain). For. Pathol. 2000, 30, 247–263. [Google Scholar] [CrossRef]
- Romero, M.A.; Sánchez, J.E.; Jiménez, J.J.; Belbahri, L.; Trapero, A.; Lefort, F.; Sanchez, M.E. New Pythium taxa causing root rot on Mediterranean Quercus species in south-west Spain and Portugal. J. Phytopathol. 2007, 155, 289–295. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Breshears, D.D.; McDowell, N. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Collins, W.; Fry, M.M.; Yu, H.; Yu, H.; Fuglestvedt, J.; Shindell, D.; West, J. Global and regional temperature-change potentials for near-term climate forcers. Atmos. Chem. Phys. 2013, 13, 2471–2485. [Google Scholar] [CrossRef] [Green Version]
- Crick, F. Central dogma of molecular biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef]
- Di Leo, A.; Claudino, W.; Colangiuli, D.; Bessi, S.; Pestrin, M.; Biganzoli, L. New strategies to identify molecular markers predicting chemotherapy activity and toxicity in breast cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2007, 18 (Suppl. S1), xii8–xii14. [Google Scholar] [CrossRef]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene expression regulation mediated through reversible m6A RNA methylation. Nat. Rev. Genet. 2014, 15, 293–306. [Google Scholar] [CrossRef]
- Srivastava, S. Emerging insights into the metabolic alterations in aging using metabolomics. Metabolites 2019, 9, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafarnia, S.; Akbarinia, M.; Hosseinpour, B.; Sanavi, S.A.M.M.; Salami, S.A. Effect of drought stress on some growth, morphological, physiological, and biochemical parameters of two different populations of Quercus brantii. iForest Biogeosci. For. 2018, 11, 212–220. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Landi, M.; Massai, R.; Nali, C.; Guidi, L.; Lorenzini, G. Variations in physiological and biochemical traits of oak seedlings grown under drought and ozone stress. Physiol. Plant. 2016, 157, 69–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Guidi, L.; Lorenzini, G.; Massai, R.; Nali, C.; Landi, M. Cross-talk between physiological and metabolic adjustments adopted by Quercus cerris to mitigate the effects of severe drought and realistic future ozone concentrations. Forests 2017, 8, 148. [Google Scholar] [CrossRef] [Green Version]
- García-Plazaola, J.L.; Hernández, A.; Fernández-Marín, B.; Esteban, R.; Peguero-Pina, J.J.; Verhoeven, A.; Cavender-Bares, J. Photoprotective mechanisms in the Genus Quercus in response to winter cold and summer drought in oaks physiological ecology. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L. Tree Physiology; Gil-Pelegrín, E., Peguero-Pina, J., Sancho-Knapik, D., Eds.; Springer: Cham, Switzerland, 2017; Volume 7, pp. 361–391. ISBN 978-3-319-69098-8. [Google Scholar]
- Arena, C.; Santorufo, L.; Cataletto, P.R.; Memoli, V.; Scudiero, R.; Maisto, G. Eco-physiological and antioxidant responses of Holm oak (Quercus ilex L.) leaves to Cd and Pb. Water Air Soil Pollut. 2017, 228, 459. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Pellegrini, E.; Guidi, L.; Landi, M.; Lorenzini, G.; Massai, R.; Remorini, D.; Tonelli, M.; Trivellini, A.; Vernieri, P.; et al. Losing the warning signal: Drought compromises the cross-talk of signaling molecules in Quercus ilex exposed to ozone. Front. Plant Sci. 2017, 8, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peguero-Pina, J.J.; Mendoza-Herrer, O.; Gil-Pelegrín, E.; Sancho-Knapik, D. Cavitation limits the recovery of gas exchange after severe drought stress in Holm Oak (Quercus ilex L.). Forest 2018, 9, 443. [Google Scholar] [CrossRef] [Green Version]
- San-Eufrasio, B.; Sánchez-Lucas, R.; López-Hidalgo, C.; Guerrero-Sanchez, V.M.; Castillejo, M.; Maldonado-Alconada, A.M.; Jorrín-Novo, J.V.; Rey, M.-D. Responses and differences in tolerance to water shortage under climatic dryness conditions in seedlings from Quercus spp. and Andalusian Q. ilex populations. Forests 2020, 11, 707. [Google Scholar] [CrossRef]
- Ghanbary, E.; Tabari Kouchaksaraei, M.; Zarafshar, M.; Bader, K.-F.M.; Mirabolfathy, M.; Ziaei, M. Differential physiological and biochemical responses of Quercus infectoria and Q. libani to drought and charcoal disease. Physiol. Plant. 2020, 168, 876–892. [Google Scholar] [CrossRef]
- Arab, L.; Seegmueller, S.; Kreuzwieser, J.; Eiblmeier, M.; Rennenberg, H. Atmospheric pCO(2) impacts leaf structural and physiological traits in Quercus petraea seedlings. Planta 2019, 249, 481–495. [Google Scholar] [CrossRef]
- Díaz-Guerra, L.; Llorens, L.; Bell, T.L.; Font, J.; González, J.A.; Verdaguer, D. Physiological, growth and root biochemical responses of Arbutus unedo and Quercus suber seedlings to UV radiation and water availability before and after aboveground biomass removal. Environ. Exp. Bot. 2019, 168, 103861. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, T.; Duan, D.; Yang, J.; Feng, L.; Zhao, G. Comparative analysis of the complete chloroplast genomes of five Quercus species. Front. Plant Sci. 2016, 7, 959. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chang, E.; Liu, J.; Huang, Y.; Wang, Y.; Yao, N.; Jiang, Z. Complete chloroplast genome sequence and phylogenetic analysis of Quercus bawanglingensis Huang, Li et Xing, a vulnerable oak tree in China. Forests 2019, 10, 587. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.; Liu, H.; Wu, S.; Yuan, Y.; Li, H.; Dong, J.; Liu, Z.; An, C.; Su, Z.; Li, B. Species identification of oaks (Quercus L., Fagaceae) from gene to genome. Int. J. Mol. Sci. 2019, 20, 5940. [Google Scholar] [CrossRef] [Green Version]
- Sork, V.; Fitz-Gibbon, S.; Puiu, D.; Crepeau, M.; Gugger, P.F.; Sherman, R.M.; Stevens, K.; Langley, C.; Pellegrini, M.; Salzberg, S. First draft assembly and annotation of the genome of a California Endemic oak Quercus lobata Née (Fagaceae). G3 Genes Genomes Genet. 2016, 6, 3485–3495. [Google Scholar] [CrossRef] [Green Version]
- Plomion, C.; Aury, J.; Amselem, J.; Alaeitabar, T.; Barbe, V.; Belser, C.; Bergès, H.; Bodénès, C.; Boudet, N.; Boury, C.; et al. Decoding the oak genome: Public release of sequence data, assembly, annotation and publication strategies. Mol. Ecol. Resour. 2016, 16, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Plomion, C.; Aury, J.; Amselem, J.; Leroy, T.; Murat, F.; Duplessis, S.; Faye, S.; Francillonne, N.; Labadie, K.; Provost, G.L.; et al. Oak genome reveals facets of long lifespan. Nat. Plants 2018, 4, 440–452. [Google Scholar] [CrossRef] [Green Version]
- Ramos, A.M.; Usie, A.; Barbosa, P.; Barros, P.M.; Capote, T.; Chaves, I.; Simoes, F.; Abreu, I.; Carrasquinho, I.; Faro, C.; et al. The draft genome sequence of cork oak. Sci. Data 2018, 5, 180069. [Google Scholar] [CrossRef] [PubMed]
- Bi, Q.; Li, D.; Zhao, Y.; Wang, M.; Li, Y.; Liu, X.; Wang, L.; Yu, H. Complete mitochondrial genome of Quercus variabilis (Fagales, Fagaceae). Mitochondrial DNA Part B 2019, 4, 3927–3928. [Google Scholar] [CrossRef] [Green Version]
- Nagamitsu, T.; Shimizu, H.; Aizawa, M.; Nakanishi, A. An admixture of Quercus dentata in the coastal ecotype of Q. mongolica var. crispula in northern Hokkaido and genetic and environmental effects on their traits. J. Plant Res. 2019, 132, 211–222. [Google Scholar] [CrossRef]
- Fernandez i Martí, A.; Romero-Rodríguez, C.; Navarro-Cerrillo, R.; Abril, N.; Jorrín-Novo, J.; Dodd, R. Population genetic diversity of Quercus ilex subsp. ballota (Desf.) Samp. reveals divergence in recent and evolutionary migration rates in the Spanish dehesas. Forests 2018, 9, 337. [Google Scholar] [CrossRef] [Green Version]
- Sork, V.L.; Squire, K.; Gugger, P.F.; Steele, S.E.; Levy, E.D.; Eckert, A.J. Landscape genomic analysis of candidate genes for climate adaptation in a California endemic oak, Quercus lobata. Am. J. Bot. 2016, 103, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Gugger, P.F.; Fitz-Gibbon, S.; PellEgrini, M.; Sork, V.L. Species-wide patterns of DNA methylation variation in Quercus lobata and their association with climate gradients. Mol. Ecol. 2016, 25, 1665–1680. [Google Scholar] [CrossRef]
- Browne, L.; Wright, J.W.; Fitz-Gibbon, S.; Gugger, P.F.; Sork, V.L. Adaptational lag to temperature in valley oak (Quercus lobata) can be mitigated by genome-informed assisted gene flow. Proc. Natl. Acad. Sci. USA 2019, 116, 25179–25185. [Google Scholar] [CrossRef] [Green Version]
- Meireles, J.E.; Beulke, A.; Borkowski, D.S.; Romero-Severson, J.; Cavender-Bares, J. Balancing selection maintains diversity in a cold tolerance gene in broadly distributed live oaks. Genome 2017, 60, 762–769. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Valiente, J.A.; Deacon, N.J.; Etterson, J.; Center, A.; Sparks, J.P.; Sparks, K.; Longwell, T.; Pilz, G.; Cavender-Bares, J. Natural selection and neutral evolutionary processes contribute to genetic divergence in leaf traits across a precipitation gradient in the tropical oak Quercus oleoides. Mol. Ecol. 2018, 27, 2176–2192. [Google Scholar] [CrossRef]
- Rellstab, C.; Zoller, S.; Walthert, L.; Lesur, I.; Pluess, A.R.; Graf, R.; Bodénès, C.; Sperisen, C.; Kremer, A.; Gugerli, F. Signatures of local adaptation in candidate genes of oaks (Quercus spp.) with respect to present and future climatic conditions. Mol. Ecol. 2016, 25, 5907–5924. [Google Scholar] [CrossRef]
- Truffaut, L.; Chancerel, E.; Ducousso, A.; Dupouey, J.L.; Badeau, V.; Ehrenmann, F.; Kremer, A. Fine-scale species distribution changes in a mixed oak stand over two successive generations. New Phytol. 2017, 215, 126–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lind-Riehl, J.F.; Gailing, O. Adaptive variation and introgression of a CONSTANS-like gene in North American red oaks. Forests 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Khodwekar, S.; Gailing, O. Evidence for environment-dependent introgression of adaptive genes between two red oak species with different drought adaptations. Am. J. Bot. 2017, 104, 1088–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pina-Martins, F.; Baptista, J.; Pappas, G.J.; Paulo, O.S. New insights into adaptation and population structure of cork oak using genotyping by sequencing. Glob. Change Biol. 2019, 25, 337–350. [Google Scholar] [CrossRef]
- Browne, L.; Mead, A.; Horn, C.; Chang, K.; Celikkol, Z.A.; Henriquez, C.L.; Ma, F.; Beraut, E.; Meyer, R.S.; Sork, V.L. Experimental DNA demethylation associates with changes in growth and gene expression of oak tree seedlings. G3 Genes Genomes Genet. 2020, 10, 1019–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inácio, V.; Barros, P.M.; Costa, A.; Roussado, C.; Gonçalves, E.; Costa, R.; Graça, J.; Oliveira, M.M.; Morais-Cecílio, L. Differential DNA methylation patterns are related to phellogen origin and quality of Quercus suber cork. PLoS ONE 2017, 12, e0169018. [Google Scholar] [CrossRef]
- Oney-Birol, S.; Fitz-Gibbon, S.; Chen, J.-M.; Gugger, P.F.; Sork, V.L. Assessment of shared alleles in drought-associated candidate genes among southern California white oak species (Quercus sect. Quercus). BMC Genet. 2018, 19, 88. [Google Scholar] [CrossRef]
- Guerrero-Sanchez, V.M.; Maldonado-Alconada, A.M.; Amil-Ruiz, F.; Verardi, A.; Jorrín-Novo, J.V.; Rey, M.-D. Ion Torrent and lllumina, two complementary RNA-seq platforms for constructing the Holm oak (Quercus ilex) transcriptome. PLoS ONE 2019, 14, e0210356. [Google Scholar] [CrossRef]
- Guerrero-Sanchez, V.M.; Maldonado-Alconada, A.M.; Amil-Ruiz, F.; Jorrin-Novo, J. V Holm oak (Quercus ilex) transcriptome. De novo sequencing and assembly analysis. Front. Mol. Biosci. 2017, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, A.; Morcuende, D.; Solla, A.; Moreno, G.; Pulido, F.; Quesada, A. Regulation by biotic stress of tannins biosynthesis in Quercus ilex: Crosstalk between defoliation and Phytophthora cinnamomi infection. Physiol. Plant. 2019, 165, 319–329. [Google Scholar] [CrossRef]
- Natali, L.; Vangelisti, A.; Guidi, L.; Remorini, D.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E.; Trivellini, A.; Vernieri, P.; et al. How Quercus ilex L. saplings face combined salt and ozone stress: A transcriptome analysis. BMC Genom. 2018, 19, 872. [Google Scholar] [CrossRef] [Green Version]
- Kotrade, P.; Sehr, E.M.; Wischnitzki, E.; Brüggemann, W. Comparative transcriptomics-based selection of suitable reference genes for normalization of RT-qPCR experiments in drought-stressed leaves of three European Quercus species. Tree Genet. Genomes 2019, 15, 38. [Google Scholar] [CrossRef]
- Madritsch, S.; Wischnitzki, E.; Kotrade, P.; Ashoub, A.; Burg, A.; Fluch, S.; Brüggemann, W.; Sehr, E.M. Elucidating drought stress tolerance in European oaks through cross-species transcriptomics. G3 Genes Genomes Genet. 2019, 9, 3181–3199. [Google Scholar] [CrossRef] [Green Version]
- Gugger, P.F.; Peñaloza-Ramírez, J.M.; Wright, J.W.; Sork, V.L. Whole-transcriptome response to water stress in a California endemic oak, Quercus lobata. Tree Physiol. 2017, 37, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Mead, A.; Peñaloza Ramirez, J.; Bartlett, M.K.; Wright, J.W.; Sack, L.; Sork, V.L. Seedling response to water stress in valley oak (Quercus lobata) is shaped by different gene networks across populations. Mol. Ecol. 2019, 28, 5248–5264. [Google Scholar] [CrossRef] [PubMed]
- Le Provost, G.; Lesur, I.; Lalanne, C.; Da Silva, C.; Labadie, K.; Aury, J.M.; Leple, J.C.; Plomion, C. Implication of the suberin pathway in adaptation to waterlogging and hypertrophied lenticels formation in pedunculate oak (Quercus robur L.). Tree Physiol. 2016, 36, 1330–1342. [Google Scholar]
- Soltani, N.; Best, T.; Grace, D.; Nelms, C.; Shumaker, K.; Romero-Severson, J.; Moses, D.; Schuster, S.; Staton, M.; Carlson, J.; et al. Transcriptome profiles of Quercus rubra responding to increased O(3) stress. BMC Genom. 2020, 21, 160. [Google Scholar] [CrossRef]
- Magalhães, A.P.; Verde, N.; Reis, F.; Martins, I.; Costa, D.; Lino-Neto, T.; Castro, P.H.; Tavares, R.M.; Azevedo, H. RNA-seq and gene network analysis uncover activation of an ABA-dependent signalosome during the cork oak root response to drought. Front. Plant Sci. 2016, 6, 1195. [Google Scholar] [CrossRef] [Green Version]
- Simova-Stoilova, L.P.; López-Hidalgo, C.; Sanchez-Lucas, R.; Valero-Galvan, J.; Romero-Rodríguez, C.; Jorrin-Novo, J.V. Holm oak proteomic response to water limitation at seedling establishment stage reveals specific changes in different plant parts as well as interaction between roots and cotyledons. Plant Sci. 2018, 276, 1–13. [Google Scholar] [CrossRef]
- Sebastiana, M.; Martins, J.; Figueiredo, A.; Monteiro, F.; Sardans, J.; Peñuelas, J.; Silva, A.; Roepstorff, P.; Pais, M.; Coelho, A. Oak protein profile alterations upon root colonization by an ectomycorrhizal fungus. Mycorrhiza 2016, 27, 109–128. [Google Scholar] [CrossRef] [Green Version]
- Suseela, V.; Tharayil, N.; Orr, G.; Hu, D. Chemical plasticity in the fine root construct of Quercus spp. varies with root order and drought. New Phytol. 2020, 228, 1835–1851. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Ubach, A.; Barbeta, A.; Sardans, J.; Guenther, A.; Ogaya, R.; Oravec, M.; Urban, O.; Peñuelas, J. Topsoil depth substantially influences the responses to drought of the foliar metabolomes of Mediterranean forests. Perspect. Plant Ecol. Evol. Syst. 2016, 21, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Gargallo-Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Peñuelas, J. Root exudate metabolomes change under drought and show limited capacity for recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar] [CrossRef] [Green Version]
- Mu, Z.; Llusià, J.; Liu, D.; Ogaya, R.; Asensio, D.; Zhang, C.; Peñuelas, J. Seasonal and diurnal variations of plant isoprenoid emissions from two dominant species in Mediterranean shrubland and forest submitted to experimental drought. Atmos. Environ. 2018, 191, 105–115. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Rodrigues, A.M.; Perdiguero, P.; António, C.; Atkin, O.K.; Li, M.; Collada, C.; Gil, L. A molecular approach to drought-induced reduction in leaf CO(2) exchange in drought-resistant Quercus ilex. Physiol. Pantarum 2018, 162, 394–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberstroh, S.; Kreuzwieser, J.; Lobo-do-Vale, R.; Caldeira, M.C.; Dubbert, M.; Werner, C. Terpenoid emissions of two Mediterranean woody species in response to drought stress. Front. Plant Sci. 2018, 9, 1071. [Google Scholar] [CrossRef] [Green Version]
- Almeida, T.; Pinto, G.; Correia, B.; Gonçalves, S.; Meijón, M.; Escandón, M. In-depth analysis of the Quercus suber metabolome under drought stress and recovery reveals potential key metabolic players. Plant Sci. 2020, 299, 110606. [Google Scholar] [CrossRef]
- Sánchez-Osorio, I.; López-Pantoja, G.; Tapias, R.; Pareja-Sánchez, E.; Domínguez, L. Monoterpene emission of Quercus suber L. highly infested by Cerambyx welensii Küster. Ann. For. Sci. 2019, 76, 98. [Google Scholar] [CrossRef]
- Saunier, A.; Ormeño, E.; Wortham, H.; Temime-Roussel, B.; Lecareux, C.; Boissard, C.; Fernandez, C. Chronic drought decreases anabolic and catabolic BVOC emissions of Quercus pubescens in a Mediterranean forest. Front. Plant Sci. 2017, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Genard-Zielinski, A.-C.; Boissard, C.; Ormeño, E.; Lathière, J.; Reiter, I.M.; Wortham, H.; Orts, J.-P.; Temime-Roussel, B.; Guenet, B.; Bartsch, S.; et al. Seasonal variations of Quercus pubescens isoprene emissions from an in natura forest under drought stress and sensitivity to future climate change in the Mediterranean area. Biogeosciences 2018, 15, 4711–4730. [Google Scholar] [CrossRef]
- Aranda, I.; Cadahía, E.; Fernández de Simón, B. Leaf ecophysiological and metabolic response in Quercus pyrenaica Willd seedlings to moderate drought under enriched CO2 atmosphere. J. Plant Physiol. 2020, 244, 153083. [Google Scholar] [CrossRef]
- Top, S.M.; Preston, C.M.; Dukes, J.S.; Tharayil, N. Climate influences the content and chemical composition of foliar tannins in green and senesced tissues of Quercus rubra. Front. Plant Sci. 2017, 8, 423. [Google Scholar] [CrossRef] [Green Version]
- Volf, M.; Weinhold, A.; Seifert, C.L.; Holicová, T.; Uthe, H.; Alander, E.; Richter, R.; Salminen, J.-P.; Wirth, C.; van Dam, N.M. Branch-localized induction promotes efficacy of volatile defences and herbivore predation in trees. J. Chem. Ecol. 2021, 47, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Contran, N.; Günthardt-Goerg, M.S.; Kuster, T.M.; Cerana, R.; Crosti, P.; Paoletti, E. Physiological and biochemical responses of Quercus pubescens to air warming and drought on acidic and calcareous soils. Plant Biol. 2013, 15 (Suppl. S1), 157–168. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Valiente, J.A.; Koehler, K.; Cavender-Bares, J. Climatic origins predict variation in photoprotective leaf pigments in response to drought and low temperatures in live oaks (Quercus series Virentes). Tree Physiol. 2015, 35, 521–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peguero-Pina, J.J.; Sancho-Knapik, D.; Morales, F.; Flexas, J.; Gil-Pelegrin, E. Differential photosynthetic performance and photoprotection mechanisms of three Mediterranean evergreen oaks under severe drought stress. Funct. Plant Biol. 2009, 36, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Ebadzad, G.; Medeira, C.; Maia, I.; Martins, J.; Cravador, A. Induction of defence responses by cinnamomins against Phytophthora cinnamomi in Quercus suber and Quercus ilex subs. rotundifolia. Eur. J. Plant Pathol. 2015, 143, 705–723. [Google Scholar] [CrossRef] [Green Version]
- Nogués, I.; Llusià, J.; Ogaya, R.; Munné-Bosch, S.; Sardans, J.; Peñuelas, J.; Loreto, F. Physiological and antioxidant responses of Quercus ilex to drought in two different seasons. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2014, 148, 268–278. [Google Scholar]
- Loewenstein, N.J.; Pallardy, S.G. Influence of a drying cycle on post-drought xylem sap abscisic acid and stomatal responses in young temperate deciduous angiosperms. New Phytol. 2002, 156, 351–361. [Google Scholar] [CrossRef]
- Hamrick, J.; Godt, M.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Gupta, P.; Varshney, R.; Sharma, P.C.; Ramesh, B. Molecular markers and their applications in wheat breeding. Plant Breed. 1999, 118, 369–390. [Google Scholar] [CrossRef]
- Müller, M.; Gailing, O. Abiotic genetic adaptation in the Fagaceae. Plant Biol. 2019, 21, 783–795. [Google Scholar] [CrossRef]
- Homolka, A.; Schueler, S.; Burg, K.; Fluch, S.; Kremer, A. Insights into drought adaptation of two European oak species revealed by nucleotide diversity of candidate genes. Tree Genet. Genomes 2013, 9, 1179–1192. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.; Gómez, F.R.; Cabrera-Puerto, R.J.; Sánchez-Cuesta, R.; Rodríguez, G.; Pérez, J. Growth and physiological sapling responses of eleven Quercus ilex ecotypes under identical environmental conditions. For. Ecol. Manag. 2018, 58–69. [Google Scholar] [CrossRef]
- Du, F.; Wang, T.; Wang, Y.; Ueno, S.; de Lafontaine, G. Contrasted patterns of local adaptation to climate change across the range of an evergreen oak, Quercus aquifolioides. Evol. Appl. 2020, 13, 2377–2391. [Google Scholar] [CrossRef]
- Modesto, I.S.; Miguel, C.; Pina-Martins, F.; Glushkova, M.; Veloso, M.; Paulo, O.; Batista, D. Identifying signatures of natural selection in cork oak (Quercus suber L.) genes through SNP analysis. Tree Genet. Genomes 2014, 10, 1645–1660. [Google Scholar] [CrossRef]
- Porth, I.; Scotti-Saintagne, C.; Barreneche, T.; Kremer, A.; Burg, K. Linkage mapping of osmotic stress induced genes of oak. Tree Genet. Genomes 2005, 1, 31–40. [Google Scholar] [CrossRef]
- Shamari, A.-R.; Mehrabi, A.-A.; Maleki, A.; Rostami, A. Association analysis of tolerance to dieback phenomena and trunk form using ISSR markers in Quercus brantii. Cell. Mol. Biol. 2018, 64, 116–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makela, M.; Michael, P.; Thériault, G.; Nkongolo, K. High genetic variation among closely related red oak (Quercus rubra) populations in an ecosystem under metal stress: Analysis of gene regulation. Genes Genom. 2016, 38, 967–976. [Google Scholar] [CrossRef]
- Casasoli, M.; Derory, J.; Morera-Dutrey, C.; Brendel, O.; Porth, I.; Guehl, J.-M.; Villani, F.; Kremer, A. Comparison of quantitative trait loci for adaptive traits between oak and chestnut based on an expressed sequence tag consensus map. Genetics 2006, 172, 533–546. [Google Scholar] [CrossRef] [Green Version]
- Parelle, J.; Zapater, M.; Scotti-Saintagne, C.; Kremer, A.; Jolivet, Y.; Dreyer, E.; Brendel, O. Quantitative trait loci of tolerance to waterlogging in a European oak (Quercus robur L.): Physiological relevance and temporal effect patterns. Plant. Cell Environ. 2007, 30, 422–434. [Google Scholar] [CrossRef]
- Brendel, O.; Le Thiec, D.; Scotti, C.; Bodénès, C.; Kremer, A.; Guehl, J. Quantitative trait loci controlling water use efficiency and related traits in Quercus robur L. Tree Genet. Genomes 2008, 4, 263–278. [Google Scholar] [CrossRef]
- Wang, L.; Du, H.; Li, T.; Wuyun, T. De novo transcriptome sequencing and identification of genes related to salt stress in Eucommia ulmoides Oliver. Trees 2018, 32, 151–163. [Google Scholar] [CrossRef]
- Ranade, S.S.; Ganea, L.-S.; Razzak, A.M.; García Gil, M.R. Fungal infection increases the rate of somatic mutation in Scots pine (Pinus sylvestris L.). J. Hered. 2015, 106, 386–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.-C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birol, I.; Raymond, A.; Jackman, S.D.; Pleasance, S.; Coope, R.; Taylor, G.A.; Yuen, M.M.S.; Keeling, C.I.; Brand, D.; Vandervalk, B.P.; et al. Assembling the 20 Gb white spruce (Picea glauca) genome from whole-genome shotgun sequencing data. Bioinformatics 2013, 29, 1492–1497. [Google Scholar] [CrossRef]
- Myburg, A.; Grattapaglia, D.; Tuskan, G.; Hellsten, U.; Hayes, R.D.; Grimwood, J.; Jenkins, J.; Lindquist, E.; Tice, H.; Bauer, D.; et al. The genome of Eucalyptus grandis. Nature 2014, 510, 356–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neale, D.B.; Wegrzyn, J.L.; Stevens, K.A.; Zimin, A.V.; Puiu, D.; Crepeau, M.W.; Cardeno, C.; Koriabine, M.; Holtz-Morris, A.E.; Liechty, J.D.; et al. Decoding the massive genome of loblolly pine using haploid DNA and novel assembly strategies. Genome Biol. 2014, 15, R59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sow, M.D.; Allona, I.; Ambroise, C.; Conde, D.; Fichot, R.; Gribkova, S.; Jorge, V.; Le-Provost, G.; Pâques, L.; Plomion, C.; et al. Epigenetics in forest trees: State of the art and potential implications for breeding and management in a context of climate change. In Plant Epigenetics Coming of Age for Breeding Applications; Mirouze, M., Bucher, E., Gallusci, P.B.T.-A., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 88, pp. 387–453. ISBN 0065-2296. [Google Scholar]
- Wegrzyn, J.; Falk, T.; Grau, E.; Buehler, S.; Ramnath, R.; Herndon, N. Cyberinfrastructure and resources to enable an integrative approach to studying forest trees. Evol. Appl. 2020, 13, 228–241. [Google Scholar] [CrossRef] [Green Version]
- Olszewska, M.; Osiecka, R. Relationship between 2C DNA content, systematic position and level of DNA endoreplication during differentiation of root parenchyma in dicot shrubs and trees-comparison with herbaceous sp. Biochem. Physiol. Pflanz. 1984, 179, 641–657. [Google Scholar] [CrossRef]
- Zoldoš, V.; Papeš, D.; Brown, S.; Panaud, O.; Šiljak-Yakovlev, S. Genome size and base composition of seven Quercus species: Inter- and intra-population variation. Genome 1998, 41, 162–168. [Google Scholar] [CrossRef]
- Kremer, A.; Casasoli, M.; Barreneche, T.; Bodenes, C.; Sisco, P.; Kubisiak, T.; Scalfi, M.; Leonardi, S.; Bakker, E.; Buiteveld, J.; et al. Fagaceae Trees. In Genome Mapping and Molecular Breeding in Plants; Springer: Cham, Switzerland, 2007; Volume 7, pp. 161–187. [Google Scholar]
- Favre, J.; Brown, S. A flow cytometric evaluation of the nuclear DNA content and GC percent in genomes of European oak species. Ann. For. Sci. 1996, 53, 915–917. [Google Scholar] [CrossRef] [Green Version]
- Rey, M.-D.; Castillejo, M.Á.; Sánchez-Lucas, R.; Guerrero-Sanchez, V.M.; López-Hidalgo, C.; Romero-Rodríguez, C.; Valero-Galván, J.; Sghaier-Hammami, B.; Simova-Stoilova, L.; Echevarría-Zomeño, S.; et al. Proteomics, holm oak (Quercus ilex L.) and other recalcitrant and orphan forest tree species: How do they see each other? Int. J. Mol. Sci. 2019, 20, 692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoldoš, V.; Papeš, D.; Cerbah, M.; Panaud, O.; Besendorfer, V.; Šiljak-yakovlev, S. Molecular-cytogenetic studies of ribosomal genes and heterochromatin reveal conserved genome organization among 11 Quercus species. Theor. Appl. Genet. 1999, 99, 969–977. [Google Scholar] [CrossRef]
- Ribeiro, T.; Loureiro, J.; Santos, C.; Morais-Cecílio, L. Evolution of rDNA FISH patterns in the Fagaceae. Tree Genet. Genomes 2011, 7, 1113–1122. [Google Scholar] [CrossRef]
- Alexander, L.W.; Woeste, K.E. Pyrosequencing of the northern red oak (Quercus rubra L.) chloroplast genome reveals high quality polymorphisms for population management. Tree Genet. Genomes 2014, 10, 803–812. [Google Scholar] [CrossRef]
- Hu, H.-L.; Zhang, J.-Y.; Li, Y.-P.; Xie, L.; Chen, D.-B.; Li, Q.; Liu, Y.-Q.; Hui, S.-R.; Qin, L. The complete chloroplast genome of the daimyo oak, Quercus dentata Thunb. Conserv. Genet. Resour. 2019, 11, 409–411. [Google Scholar] [CrossRef]
- Hu, H.-L.; Wang, L.-Z.; Yang, J.; Zhang, R.-S.; Li, Q.; Liu, Y.-Q.; Qin, L. The complete chloroplast genome of Quercus fenchengensis and the phylogenetic implication. Mitochondrial DNA Part B 2019, 4, 3066–3067. [Google Scholar] [CrossRef] [PubMed]
- Harfouche, A.; Meilan, R.; Altman, A. Molecular and physiological responses to abiotic stress in forest trees and their relevance to tree improvement. Tree Physiol. 2014, 34, 1181–1198. [Google Scholar] [CrossRef] [Green Version]
- Della Coletta, R.; Qiu, Y.; Ou, S.; Hufford, M.B.; Hirsch, C.N. How the pan-genome is changing crop genomics and improvement. Genome Biol. 2021, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and promises of epigenetics for forest trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Gutzat, R.; Scheid, O.M. Epigenetic responses to stress: Triple defense? Curr. Opin. Plant Biol. 2012, 15, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Turgut-Kara, N.; As1kan, B.; Celik, H. Epigenetic memory and priming in plants. Genetica 2020, 148, 47–54. [Google Scholar] [CrossRef]
- Law, J.A.; Jacobsen, S.E. Molecular biology. Dynamic DNA methylation. Science 2009, 323, 1568–1569. [Google Scholar] [CrossRef]
- Johannes, F.; Schmitz, R.J. Spontaneous epimutations in plants. New Phytol. 2019, 221, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Correia, B.; Valledor, L.; Meijón, M.; Rodriguez, J.L.; Dias, M.C.; Santos, C.; Cañal, M.J.; Rodriguez, R.; Pinto, G. Is the interplay between epigenetic markers related to the acclimation of cork oak plants to high temperatures? PLoS ONE 2013, 8, e53543. [Google Scholar] [CrossRef] [Green Version]
- Rico, L.; Ogaya, R.; Barbeta, A.; Penuelas, J. Changes in DNA methylation fingerprint of Quercus ilex trees in response to experimental field drought simulating projected climate change. Plant Biol. 2014, 16, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Platt, A.; Gugger, P.F.; Pellegrini, M.; Sork, V.L. Genome-wide signature of local adaptation linked to variable CpG methylation in oak populations. Mol. Ecol. 2015, 24, 3823–3830. [Google Scholar] [CrossRef]
- Chwialkowska, K.; Korotko, U.; Kosinska, J.; Szarejko, I.; Kwasniewski, M. Methylation Sensitive Amplification Polymorphism Sequencing (MSAP-Seq)-a method for high-throughput analysis of differentially methylated CCGG sites in plants with large genomes. Front. Plant Sci. 2017, 8, 2056. [Google Scholar] [CrossRef] [Green Version]
- Espinas, N.A.; Saze, H.; Saijo, Y. Epigenetic control of defense signaling and priming in plants. Front. Plant Sci. 2016, 7, 1201. [Google Scholar] [CrossRef] [PubMed]
- Balao, F.; Paun, O.; Alonso, C. Uncovering the contribution of epigenetics to plant phenotypic variation in Mediterranean ecosystems. Plant Biol. 2018, 20 (Suppl. S1), 38–49. [Google Scholar] [CrossRef]
- Yakovlev, I.; Carneros, E.; Lee, Y.; Olsen, J.; Fossdal, C.G. Transcriptional profiling of epigenetic regulators in somatic embryos during temperature induced formation of an epigenetic memory in Norway spruce. Planta 2016, 243, 1237–1249. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Schwachtje, J.; Baier, M.; Balazadeh, S.; Bäurle, I.; Geiselhardt, S.; Hincha, D.; Kunze, R.; Mueller-Roeber, B.; Rillig, M.; et al. Priming and memory of stress responses in organisms lacking a nervous system. Biol. Rev. 2016, 91, 753–761. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant immunity: From signaling to epigenetic control of defense. Trends Plant Sci. 2018, 23, 833–844. [Google Scholar] [CrossRef]
- Carneros, E.; Yakovlev, I.; Viejo, M.; Olsen, J.E.; Fossdal, C.G. The epigenetic memory of temperature during embryogenesis modifies the expression of bud burst-related genes in Norway spruce epitypes. Planta 2017, 246, 553–566. [Google Scholar] [CrossRef]
- Avramova, Z. Defence-related priming and responses to recurring drought: Two manifestations of plant transcriptional memory mediated by the ABA and JA signalling pathways. Plant. Cell Environ. 2019, 42, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Schmülling, T. Stress priming, memory, and signalling in plants. Plant. Cell Environ. 2019, 42, 753–761. [Google Scholar] [CrossRef]
- Gallusci, P.; Dai, Z.; Génard, M.; Gauffretau, A.; Leblanc-Fournier, N.; Richard-Molard, C.; Vile, D.; Brunel-Muguet, S. Epigenetics for plant improvement: Current knowledge and modeling avenues. Trends Plant Sci. 2017, 22, 610–623. [Google Scholar] [CrossRef] [PubMed]
- Kukurba, K.R.; Montgomery, S.B. RNA sequencing and analysis. Cold Spring Harb. Protoc. 2015, 2015, 951–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spieß, N.; Oufir, M.; Matušíková, I.; Stierschneider, M.; Kopecky, D.; Homolka, A.; Burg, K.; Fluch, S.; Hausman, J.; Wilhelm, E. Ecophysiological and transcriptomic responses of oak (Quercus robur) to long-term drought exposure and rewatering. Environ. Exp. Bot. 2012, 77, 117–126. [Google Scholar] [CrossRef]
- Almeida, T.; Pinto, G.; Correia, B.; Santos, C.; Gonçalves, S. Plant Physiology and Biochemistry QsMYB1 expression is modulated in response to heat and drought stresses and during plant recovery in Quercus suber. Plant Physiol. Biochem. 2013, 73, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Rasheed-Depardieu, C.; Parelle, J.; Tatin-Froux, F.; Parent, C.; Capelli, N. Short-term response to waterlogging in Quercus petraea and Quercus robur: A study of the root hydraulic responses and the transcriptional pattern of aquaporins. Plant Physiol. Biochem. 2015, 97, 323–330. [Google Scholar] [CrossRef]
- Ebadzad, G.; Cravador, A. Quantitative RT-PCR analysis of differentially expressed genes in Quercus suber in response to Phytophthora cinnamomi infection. Springerplus 2014, 3, 613. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Ueno, S.; Le Provost, G.; Léger, V.; Klopp, C.; Noirot, C.; Frigerio, J.-M.; Salin, F.; Salse, J.; Abrouk, M.; Murat, F.; et al. Bioinformatic analysis of ESTs collected by Sanger and pyrosequencing methods for a keystone forest tree species: Oak. BMC Genom. 2010, 11, 650. [Google Scholar] [CrossRef] [Green Version]
- Tarkka, M.T.; Herrmann, S.; Wubet, T.; Feldhahn, L.; Recht, S.; Kurth, F.; Mailänder, S.; Bönn, M.; Neef, M.; Angay, O.; et al. OakContigDF159.1, a reference library for studying differential gene expression in Quercus robur during controlled biotic interactions: Use for quantitative transcriptomic profiling of oak roots in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Leal, J.B.; Abreu, I.A.; Alabaca, C.S.; Almeida, M.H.; Almeida, P.; Almeida, T.; Amorim, M.I.; Araujo, S.; Azevedo, H.; Badia, A.; et al. A comprehensive assessment of the transcriptome of cork oak (Quercus suber) through EST sequencing. BMC Genom. 2014, 15, 371. [Google Scholar] [CrossRef] [Green Version]
- López-Hidalgo, C.; Guerrero-Sánchez, V.M.; Gómez-Gálvez, I.; Sánchez-Lucas, R.; Castillejo-Sánchez, M.A.; Maldonado-Alconada, A.M.; Valledor, L.; Jorrín-Novo, J.V. A multi-omics analysis pipeline for the metabolic pathway reconstruction in the orphan species Quercus ilex. Front. Plant Sci. 2018, 9, 935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Sanchez, V.M.; Maldonado-Alconada, A.M.; Sánchez-Lucas, R.; Rey, M.D. Specific protein database creation from transcriptomics data in nonmodel species: Holm oak (Quercus ilex L.). In Plant Proteomics. Methods in Molecular Biology; Jorrin-Novo, J., Valledor, L., Castillejo, M.A., Rey, M., Eds.; Springer: New York, NY, USA, 2020; Volume 2139, pp. 57–68. ISBN 978-1-0716-0528-8. [Google Scholar]
- Park, O.K. Proteomic studies in plants. J. Biochem. Mol. Biol. 2004, 37, 133–138. [Google Scholar] [CrossRef]
- Jorge, I.; Navarro, R.M.; Lenz, C.; Ariza, D.; Jorrín, J. Variation in the Holm oak leaf proteome at different plant developmental stages, between provenances and in response to drought stress. Proteomics 2006, 6 (Suppl. S1), S207–S214. [Google Scholar] [CrossRef]
- Romero-Rodríguez, M.C.; Pascual, J.; Valledor, L.; Jorrín-Novo, J. Improving the quality of protein identification in non-model species. Characterization of Quercus ilex seed and Pinus radiata needle proteomes by using SEQUEST and custom databases. J. Proteom. 2014, 105, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Gálvez, I.; Sánchez-Lucas, R.; San-Eufrasio, B.; Rodríguez de Francisco, L.E.; Maldonado-Alconada, A.M.; Fuentes-Almagro, C.; Castillejo, M.A. Optimizing shotgun proteomics analysis for a confident protein identification and quantitation in orphan plant species: The case of Holm oak (Quercus ilex). In Plant Proteomics. Methods in Molecular Biology; Jorrin-Novo, J., Valledor, L., Castillejo, M.A., Rey, M., Eds.; Springer: New York, NY, USA, 2020; Volume 2139, pp. 157–168. ISBN 978-1-0716-0527-1. [Google Scholar]
- Escandón, M.; Jorrín-Novo, J.V.; Castillejo, M.Á. Application and optimization of label-free shotgun approaches in the study of Quercus ilex. J. Proteom. 2020, 233, 104082. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Castillejo, M.Á.; Baazaoui, N.; Jorrín-Novo, J.V.; Escandón, M. GeLC-Orbitrap/MS and 2-DE-MALDI-TOF/TOF comparative proteomics analysis of seed cotyledons from the non-orthodox Quercus ilex tree species. J. Proteom. 2021, 233, 104087. [Google Scholar] [CrossRef]
- Valero-Galván, J.; Valledor, L.; Navarro-Cerrillo, R.M.; Gil Pelegrín, E.; Jorrín-Novo, J.V. Studies of variability in Holm oak (Quercus ilex subsp. ballota [Desf.] Samp.) through acorn protein profile analysis. J. Proteom. 2011, 74, 1244–1255. [Google Scholar] [CrossRef] [PubMed]
- Valero-Galván, J.; Sghaier-Hammami, B.; Navarro-Cerrillo, R.M.; Jorrín-Novo, J.V. Natural variability and responses to stresses in andalusia Holm oak (Quercus ilex subsp. ballota) populations. In Oak: Ecology, Types and Management; Chuteira, C.A., Grão, A.B., Eds.; Nova Science Publisher: Hauppauge, NY, USA, 2012; pp. 193–206. ISBN 978-1-61942-492-0. [Google Scholar]
- Valero-Galván, J.; González, R.; Valledor, L.; Navarro-Cerrillo, R.M.; Jorrin-Novo, J.V. Proteotyping of Holm oak (Quercus ilex subsp. ballota) provenances through proteomic analysis of acorn flour. Methods Mol. Biol. 2014, 1072, 709–723. [Google Scholar]
- López-Hidalgo, C.; Escandón, M.; Valledor, L.; Jorrin-Novo, J.V. A pipeline for metabolic pathway reconstruction in plant orphan species. In Plant Proteomics. Methods in Molecular Biology; Jorrin-Novo, J., Valledor, L., Castillejo, M.A., Rey, M., Eds.; Springer: New York, NY, USA, 2020; Volume 2139, pp. 367–380. [Google Scholar]
- Romero-Rodríguez, M.C.; Abril, N.; Sánchez-Lucas, R.; Jorrín-Novo, J.V. Multiplex staining of 2-DE gels for an initial phosphoproteome analysis of germinating seeds and early grown seedlings from a non-orthodox specie: Quercus ilex L. subsp. ballota [Desf.] Samp. Front. Plant Sci. 2015, 6, 620. [Google Scholar] [CrossRef] [Green Version]
- Romero-Rodríguez, M.C.; Jorrín-Novo, J.V.; Castillejo, M.A. Toward characterizing germination and early growth in the non-orthodox forest tree species Quercus ilex through complementary gel and gel-free proteomic analysis of embryo and seedlings. J. Proteom. 2019, 197, 60–70. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Redondo-López, I.; Valero-Galvàn, J.; Jorrín-Novo, J.V. Protein profile of cotyledon, tegument, and embryonic axis of mature acorns from a non-orthodox plant species: Quercus ilex. Planta 2016, 243, 369–396. [Google Scholar] [CrossRef] [PubMed]
- Sghaier-Hammami, B.; Hammami, S.B.M.; Baazaoui, N.; Gómez-Díaz, C.; Jorrín-Novo, J.V. Dissecting the seed maturation and germination processes in the Non-Orthodox Quercus ilex species based on protein signatures as revealed by 2-DE coupled to MALDI-TOF/TOF proteomics strategy. Int. J. Mol. Sci. 2020, 21, 4870. [Google Scholar] [CrossRef] [PubMed]
- Ricardo, C.P.P.; Martins, I.; Francisco, R.; Sergeant, K.; Pinheiro, C.; Campos, A.; Renaut, J.; Fevereiro, P. Proteins associated with cork formation in Quercus suber L. stem tissues. J. Proteom. 2011, 74, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Wienkoop, S.; de Almeida, J.F.; Brunetti, C.; Zarrouk, O.; Planchon, S.; Gori, A.; Tattini, M.; Ricardo, C.P.; Renaut, J.; et al. Phellem cell-wall components are discriminants of cork quality in Quercus suber. Front. Plant Sci. 2019, 10, 944. [Google Scholar] [CrossRef] [Green Version]
- Gómez, A.; López, J.A.; Pintos, B.; Camafeita, E.; Bueno, M.A. Proteomic analysis from haploid and diploid embryos of Quercus suber L. identifies qualitative and quantitative differential expression patterns. Proteomics 2009, 9, 4355–4367. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Garay, A.; Lopez, J.A.; Camafeita, E.; Bueno, M.A.; Pintos, B. Proteomic perspective of Quercus suber somatic embryogenesis. J. Proteom. 2013, 93, 314–325. [Google Scholar] [CrossRef]
- Huerta-Ocampo, J.Á.; Valenzuela-Corral, A.; Robles-Burgueño, M.D.R.; Guzmán-Partida, A.M.; Hernández-Oñate, M.Á.; Vázquez-Moreno, L.; Pavón-Romero, G.F.; Terán, L.M. Proteomic identification of allergenic proteins in red oak (Quercus rubra) pollen. World Allergy Organ. J. 2020, 13, 100111. [Google Scholar] [CrossRef]
- Pedrosa, M.; Guerrero-Sanchez, V.M.; Canales-Bueno, N.; Loli-Ausejo, D.; Castillejo, M.Á.; Quirce, S.; Jorrin-Novo, J.V.; Rodriguez-Perez, R. Quercus ilex pollen allergen, Que i 1, responsible for pollen food allergy syndrome caused by fruits in Spanish allergic patients. Clin. Exp. Allergy 2020, 50, 815–823. [Google Scholar] [CrossRef]
- Rey, M.D.; Valledor, L.; Castillejo, M.A.; Sánchez-Lucas, R.; López-Hidalgo, C.; Guerrero-Sánchez, V.M.; Colina, F.J.; Escandón, M.; Maldonado-Alconada, A.M.; Jorrín-Novo, J.V. Recent advances in MS-based plant proteomics: Proteomics data validation through integration with other classic and -omics approaches. In Progress in Botany; Cánovas, F., Lüttge, U., Leuschner, C., Risueño, M., Eds.; Springer: Cham, Switzerland, 2019; Volume 81, ISBN 978-3-030-36327-7. [Google Scholar]
- Echevarría-Zomeño, S.; Ariza, D.; Jorge, I.; Lenz, C.; del Campo, A.D.; Jorrín, J.; Navarro, R.M. Changes in the protein profile of Quercus ilex leaves in response to drought stress and recovery. J. Plant Physiol. 2009, 166, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Valero-Galván, J.; González-Fernández, R.; Navarro-Cerrillo, R.; Gil-Pelegrin, E.; Jorrín-Novo, J. Physiological and proteomic analyses of drought stress response in Holm oak provenances. J. Proteome Res. 2013, 12, 5110–5123. [Google Scholar] [CrossRef] [PubMed]
- Simova-Stoilova, L.P.; Romero-Rodríguez, M.C.; Sánchez-Lucas, R.; Navarro-Cerrillo, R.M.; Medina-Aunon, J.A.; Jorrín-Novo, J.V. 2-DE proteomics analysis of drought treated seedlings of Quercus ilex supports a root active strategy for metabolic adaptation in response to water shortage. Front. Plant Sci. 2015, 6, 627. [Google Scholar] [CrossRef] [Green Version]
- Sghaier-Hammami, B.; Valero-Galvàn, J.; Romero-Rodríguez, M.C.; Navarro-Cerrillo, R.M.; Abdelly, C.; Jorrín-Novo, J. Physiological and proteomics analyses of Holm oak (Quercus ilex subsp. ballota [Desf.] Samp.) responses to Phytophthora cinnamomi. Plant Physiol. Biochem. 2013, 71, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, K.; Spiess, N.; Renaut, J.; Wilhelm, E.; Hausman, J.F. One dry summer: A leaf proteome study on the response of oak to drought exposure. J. Proteom. 2011, 74, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, H.; O’Gorman, A.; Brennan, L. Metabolomics as a tool in nutritional research. Curr. Opin. Lipidol. 2015, 26, 30–34. [Google Scholar] [CrossRef]
- Patel, S.; Ahmed, S. Emerging field of metabolomics: Big promise for cancer biomarker identification and drug discovery. J. Pharm. Biomed. Anal. 2015, 107, 63–74. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, F.; Deng, Y.; Zhao, Y.; Ren, H. Metagenomic and metabolomic analysis of the toxic effects of trichloroacetamide-induced gut microbiome and urine metabolome perturbations in mice. J. Proteome Res. 2015, 14, 1752–1761. [Google Scholar] [CrossRef]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef]
- Monteiro, M.S.; Carvalho, M.; Bastos, M.L.; Guedes de Pinho, P. Metabolomics analysis for biomarker discovery: Advances and challenges. Curr. Med. Chem. 2013, 20, 257–271. [Google Scholar] [CrossRef]
- Escandón, M.; Meijón, M.; Valledor, L.; Pascual, J.; Pinto, G.; Cañal, M.J. Metabolome integrated analysis of high-temperature response in Pinus radiata. Front. Plant Sci. 2018, 9, 485. [Google Scholar] [CrossRef]
- López-Hidalgo, C.; Trigueros, M.; Menéndez, M.; Jorrin-Novo, J.V. Phytochemical composition and variability in Quercus ilex acorn morphotypes as determined by NIRS and MS-based approaches. Food Chem. 2021, 338, 127803. [Google Scholar] [CrossRef]
- Schwanz, P.; Polle, A. Differential stress responses of antioxidative systems to drought in pendunculate oak (Quercus robur) and maritime pine (Pinus pinaster) grown under high CO2 concentrations. J. Exp. Bot. 2001, 52, 133–143. [Google Scholar] [PubMed]
- Chaves, I.; Passarinho, J.A.P.; Capitão, C.; Chaves, M.M.; Fevereiro, P.; Ricardo, C.P.P. Temperature stress effects in Quercus suber leaf metabolism. J. Plant Physiol. 2011, 168, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F. Distribution of isoprenoid emitters in the Quercus genus around the world: Chemo-taxonomical implications and evolutionary considerations based on the ecological function of the trait. Perspect. Plant Ecol. Evol. Syst. 2002, 5, 185–192. [Google Scholar] [CrossRef]
- Loreto, F.; Bagnoli, F.; Fineschi, S. One species, many terpenes: Matching chemical and biological diversity. Trends Plant Sci. 2009, 14, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Llusià, J. Linking photorespiration, monoterpenes and thermotolerance in Quercus. New Phytol. 2002, 155, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Ubach, A.; Gargallo-Garriga, A.; Sardans, J.; Oravec, M.; Mateu-Castell, L.; Pérez-Trujillo, M.; Parella, T.; Ogaya, R.; Urban, O.; Peñuelas, J. Drought enhances folivory by shifting foliar metabolomes in Quercus ilex trees. New Phytol. 2014, 202, 874–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, T.; Niinemets, Ü.; Sabate, S.; Gracia, C.; Peñuelas, J. Seasonality of monoterpene emission potentials in Quercus ilex and Pinus pinea: Implications for regional VOC emissions modeling. J. Geophys. Res. Atmos. 2009, 114. [Google Scholar] [CrossRef] [Green Version]
- Genard-Zielinski, A.-C.; Boissard, C.; Fernandez, C.; Kalogridis, C.; Lathière, J.; Gros, V.; Bonnaire, N.; Ormeño, E. Variability of BVOC emissions from a Mediterranean mixed forest in southern France with a focus on Quercus pubescens. Atmos. Chem. Phys. 2015, 15, 431–446. [Google Scholar] [CrossRef] [Green Version]
- Llusià, J.; Peñuelas, J.; Seco, R.; Filella, I. Seasonal changes in the daily emission rates of terpenes by Quercus ilex and the atmospheric concentrations of terpenes in the natural park of Montseny, NE Spain. J. Atmos. Chem. 2012, 69, 215–230. [Google Scholar] [CrossRef]
- Loreto, F.; Schnitzler, J.-P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Llusia, J.; Roahtyn, S.; Yakir, D.; Rotenberg, E.; Seco, R.; Guenther, A.; Peñuelas, J. Photosynthesis, stomatal conductance and terpene emission response to water availability in dry and mesic Mediterranean forests. Trees 2016, 30, 749–759. [Google Scholar] [CrossRef] [Green Version]
- Brüggemann, N.; Schnitzler, J. Comparison of isoprene emission, intercellular isoprene concentration and photosynthetic performance in water-limited oak (Quercus pubescens Willd. and Quercus robur L.) saplings. Plant Biol. 2002, 4, 456–463. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Buatois, B.; Chiche, E.; Shahin, O.; Staudt, M. Leaf isoprene emission declines in Quercus pubescens seedlings experiencing drought—Any implication of soluble sugars and mitochondrial respiration? Environ. Exp. Bot. 2013, 85, 36–42. [Google Scholar] [CrossRef]
- Llusià, J.; Peñuelas, J.; Asensio, D.; Munné-Bosch, S. Airborne limonene confers limited thermotolerance to Quercus ilex. Physiol. Plant. 2005, 123, 40–48. [Google Scholar] [CrossRef]
- Lavoir, A.-V.; Staudt, M.; Schnitzler, J.; Landais, D.; Massol, F.; Rocheteau, A.; Rodríguez, R.; Zimmer, I.; Rambal, S. Drought reduced monoterpene emissions from the evergreen Mediterranean oak Quercus ilex: Results from a throughfall displacement experiment. Biogeosciences 2009, 6, 1167–1180. [Google Scholar] [CrossRef] [Green Version]
- Bourtsoukidis, E.; Kawaletz, H.; Radacki, D.; Schütz, S.; Hakola, H.; Hellén, H.; Noe, S.; Mölder, I.; Ammer, C.; Bonn, B. Impact of flooding and drought conditions on the emission of volatile organic compounds of Quercus robur and Prunus serotina. Trees 2014, 28, 193–204. [Google Scholar] [CrossRef]
- Faiola, C.; Taipale, D. Impact of insect herbivory on plant stress volatile emissions from trees: A synthesis of quantitative measurements and recommendations for future research. Atmos. Environ. X 2020, 5, 100060. [Google Scholar] [CrossRef]
- Suseela, V.; Tharayil, N.; Xing, B.; Dukes, J.S. Warming and drought differentially influence the production and resorption of elemental and metabolic nitrogen pools in Quercus rubra. Glob. Change Biol. 2015, 21, 4177–4195. [Google Scholar] [CrossRef] [PubMed]
- Palsson, B. Systems Biology. Constraint-Based Reconstruction and Analysis; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Breitling, R. What is systems biology? Front. Physiol. 2010, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Escandón, M.; Valledor, L.; Pascual, J.; Pinto, G.; Cañal, M.J.; Meijón, M. System-wide analysis of short-term response to high temperature in Pinus radiata. J. Exp. Bot. 2017, 68, 3629–3641. [Google Scholar] [CrossRef]
- Pascual, J.; Cañal, M.J.; Escandón, M.; Meijón, M.; Weckwerth, W.; Valledor, L. Integrated physiological, proteomic, and metabolomic analysis of Ultra Violet (UV) stress responses and adaptation mechanisms in Pinus radiata. MCP 2017, 16, 485–501. [Google Scholar] [PubMed] [Green Version]
- Meijón, M.; Feito, I.; Oravec, M.; Delatorre, C.; Weckwerth, W.; Majada, J.; Valledor, L. Exploring natural variation of Pinus pinaster Aiton using metabolomics: Is it possible to identify the region of origin of a pine from its metabolites? Mol. Ecol. 2016, 25, 959–976. [Google Scholar] [CrossRef] [PubMed]
- Kremer, A.; Abbott, A.G.; Carlson, J.E.; Manos, P.S.; Plomion, C.; Sisco, P.; Staton, M.E.; Ueno, S.; Vendramin, G.G. Genomics of Fagaceae. Tree Genet. Genomes 2012, 8, 583–610. [Google Scholar] [CrossRef] [Green Version]
- Initiative, T.A.G. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estravis-Barcala, M.; Mattera, M.G.; Soliani, C.; Bellora, N.; Opgenoorth, L.; Heer, K.; Arana, M.V. Molecular bases of responses to abiotic stress in trees. J. Exp. Bot. 2020, 71, 3765–3779. [Google Scholar] [CrossRef]
- Weighill, D.; Tschaplinski, T.J.; Tuskan, G.A.; Jacobson, D. Data Integration in Poplar: ‘Omics Layers and Integration Strategies. Front. Genet. 2019, 10, 874. [Google Scholar] [CrossRef]
- Martínez, M.T.; Corredoira, E.; Vieitez, A.; Cernadas, M.J.; Montenegro, R.; Ballester, A.; Vieitez, F.J.; José, M.C.S. Micropropagation of mature Quercus ilex L. trees by axillary budding. Plant Cell Tissue Organ Cult. 2017, 131, 499–512. [Google Scholar] [CrossRef]
- Plomion, C.; Bastien, C.; Bogeat-Triboulot, M.-B.; Bouffier, L.; Déjardin, A.; Duplessis, S.; Fady, B.; Heuertz, M.; Gac, A.; Provost, G.; et al. Forest tree genomics: 10 achievements from the past 10 years and future prospects. Ann. For. Sci. 2016, 73, 77–103. [Google Scholar] [CrossRef] [Green Version]


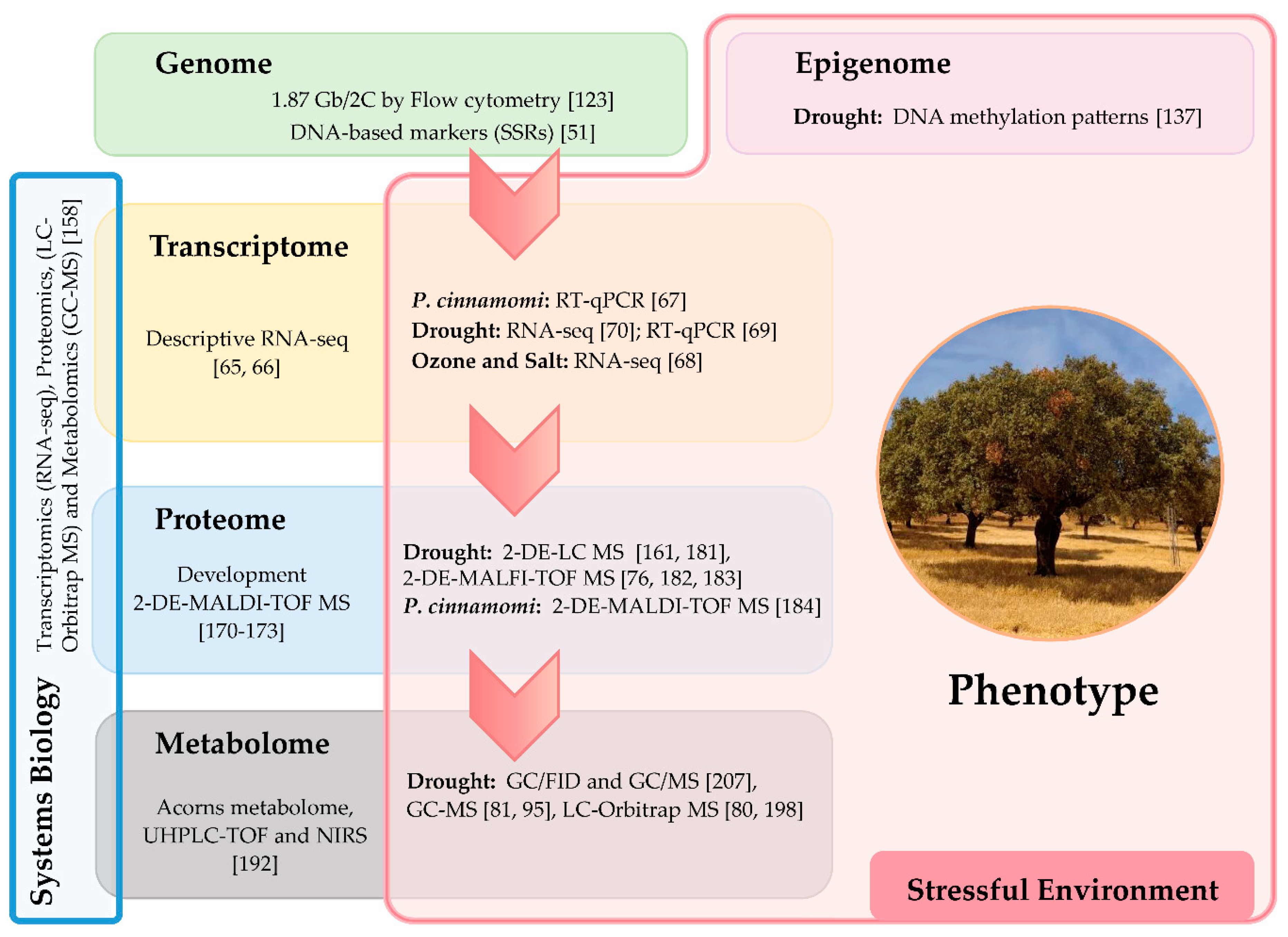{kind=link}
| Habitus | Technique/Assay | Abiotic or Biotic Stress | Reference | |
|---|---|---|---|---|
| Classical Biochemistry | ||||
| Q. brantii | Deciduous | Pigments, proline, total phenolic and flavonoids content, lipid peroxidation, ROS determination, antioxidant enzymes and PAL activity | Drought | Jafarnia et al. [31] |
| Q. cerris | Deciduous | Pigments and proline content, lipid peroxidation | Drought and O3 | Cotrozzi et al. [32] |
| Pigments and proline content, lipid peroxidation, sugar and ABA content | Drought and O3 | Cotrozzi et al. [33] | ||
| Q. coccifera | Evergreen | Pigments content | Drought and cold | García-Plazaola et al. [34] |
| Q. ilex | Evergreen | Pigments and proline content, lipid peroxidation | Drought and O3 | Cotrozzi et al. [32] |
| Pigments content | Drought and cold | García-Plazaola et al. [34] | ||
| Pigments content, ROS determination and antioxidant enzymes activity | Soil pollutants (Pb and Cd) | Arena et al. [35] | ||
| ROS, proline and phytohormones determination | Drought and O3 | Cotrozzi et al. [36] | ||
| ABA determination | Drought | Peguero-Pina et al. [37] | ||
| Pigments, sugar, total phenolic and amino acid content | Drought | San-Eufrasio et al. [38] | ||
| Q. infectoria | Semi evergreen | Proline, sugar, total phenolic and flavonoids content, lipid peroxidation, ROS determination, antioxidant enzymes and PAL activity | Drought and charcoal disease | Ghanbary et al. [39] |
| Q. libani | Deciduous | Proline, sugar, total phenolic and flavonoids content, lipid peroxidation, ROS determination, antioxidant enzymes and PAL activity | Drought and charcoal disease | Ghanbary et al. [39] |
| Q. pubescens | Deciduous | Pigments and proline content, lipid peroxidation | Drought and O3 | Cotrozzi et al. [32] |
| Pigments content | Drought and cold | García-Plazaola et al. [34] | ||
| Q. petraea | Deciduous | Lignin, sugar, cellulose, lipid peroxidation and amino acids content, ROS determination, and GR activity | Partial pressure pCO2 | Arab et al. [40] |
| Q. suber | Evergreen | Starch, sugar, total phenolic content | Drought and UV radiation | Diaz-Guerra et al. [41] |
| Genome | ||||
| Q. aliena | Deciduous | Chloroplast genome sequencing | - | Yang et al. [42] |
| Q. aliena var. acuteserrata | Deciduous | Chloroplast genome sequencing | - | Yang et al. [42] |
| Q. baronii | Evergreen | Chloroplast genome sequencing | - | Yang et al. [42] |
| Q. bawanglingensis | Evergreen | Chloroplast genome sequencing | - | Liu et al. [43] |
| Q. dentata | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. dolicholepis | Evergreen | Chloroplast genome sequencing | - | Yang et al. [42] |
| Q. fabri | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. gambelii | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. glandulifera var. brevipetiolata | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. lobata | Deciduous | Whole genome sequencing | - | Sork et al. [45] |
| Q. macrocarpa | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. palustris | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. phillyraeoides | Evergreen | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. robur | Deciduous | Whole genome sequencing | - | Plomion et al. [46,47] |
| Q. rubra | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. serrata | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. stellata | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| Q. suber | Evergreen | Whole genome sequencing | - | Ramos et al. [48] |
| Q. variabilis | Deciduous | Mitochondrial and chloroplast genome sequencing | - | Yang et al. [42]; Bi et al. [49]; Pang et al. [44] |
| Q. wutaishanica | Deciduous | Chloroplast genome sequencing | - | Pang et al. [44] |
| DNA Based Markers | ||||
| Q. dentata | Deciduous | SSR | Climate adaptation | Nagamitsu et al. [50] |
| Q. ilex | Evergreen | SSR | Climate adaptation | Fernandez i Marti et al. [51] |
| Q. lobata | Deciduous | SNP | Climate adaptation | Sork et al. [52]; Gugger et al. [53]; Browne et al. [54] |
| Q. mongolica | Deciduous | SSR | Climate adaptation | Nagamitsu et al. [50] |
| Q. oleoides | Evergreen | SNP | Cold | Meireles et al. [55] |
| SSR | Drought | Ramírez-Valiente et al. [56] | ||
| Q. petraea | Deciduous | SNP | Climate adaptation | Rellstab et al. [57] |
| SNP | Climate adaptation | Truffaut et al. [58] | ||
| Q. robur | Deciduous | SNP | Climate adaptation | Truffaut et al. [58] |
| Q. rubra | Deciduous | SSR | Drought | Lind-Riehl and Gailing, [59]; Khodwekar and Gailing [60] |
| Q. suber | Evergreen | SNP | Climate adaptation | Pina-Martins et al. [61] |
| Epigenome | ||||
| Q. lobata | Deciduous | Single methylation variants | Climate gradients | Gugger et al. [53] |
| DNA Demethylation by 5-Azacytidine | Climate adaptation | Browne et al. [62] | ||
| Q. suber | Evergreen | Single methylation variants | Climate adaptation | Inácio et al. [63] |
| Transcriptome | ||||
| Q. berberidifolia | Evergreen | RNA-Seq | Drought | Oney-Birol et al. [64] |
| Q. cornelius-mulleri | Evergreen | RNA-Seq | Drought | Oney-Birol et al. [64] |
| Q. engelmannii, | Semi evergreen | RNA-Seq | Drought | Oney-Birol et al. [64] |
| Q. ilex | Evergreen | RNA-Seq | - | Guerrero-Sánchez et al. [65,66] |
| RT-qPCR | Phytophthora cinnamomi | Gallardo et al. [67] | ||
| RNA-Seq | Salt and O3 | Natali et al. [68] | ||
| RT-qPCR | Drought | Kotrade et al. [69] | ||
| RNA-Seq | Drought | Madritsch et al. [70] | ||
| Q. lobata | Deciduous | RNA-Seq | Drought | Gugger et al. [71]; Mead et al. [72] |
| Q. pubescens | Deciduous | RNA-Seq | Drought | Kotrade et al. [69]; Madritsch et al. [70] |
| Q. petraea | Deciduous | RNA-Seq | Waterlogging | Le Provost et al. [73] |
| Q. robur | Deciduous | RNA-Seq | Waterlogging | Le Provost et al. [73] |
| RT-qPCR | Drought | Kotrade et al. [69] | ||
| RNA-Seq | Drought | Madritsch et al. [70] | ||
| Q. rubra | Deciduous | RNA-Seq | O3 | Soltani et al. [74] |
| Q. suber | Evergreen | RNA-Seq | Drought | Magalhães et al. [75] |
| Proteome | ||||
| Q. ilex | Evergreen | 2-DE MALDI TOF/TOF | Drought | Simova-Stoilova et al. [76] |
| Q. suber | Evergreen | 2D-DIGE MALDI-TOF/TOF | Ectomycorrhizal | Sebastiana et al. [77] |
| Metabolome | ||||
| Q. alba | Deciduous | HPLC-Orbitrap-MS | Drought | Suseela et al. [78] |
| Q. ilex | Evergreen | LC-Orbitrap-MS | Drought | Rivas-Ubach et al. [79]; Gargallo-Garriga et al. [80] |
| GC-MS | Drought | Mu et al. [81]; Rodríguez-Calcerrada et al. [82] | ||
| Q. suber | Evergreen | GC-MS | Drought | Haberstroh et al. [83] |
| LC-Orbitrap-MS | Drought | Almeida et al. [84] | ||
| GC-MS | Cerambyx welensii | Sánchez-Osorio et al. [85] | ||
| Q. pubescens | Deciduous | PTR-TOF-MS | Drought | Saunier et al. [86] |
| PTR-MS/ GC-MS | Drought | Genard-Zielinski et al. [87] | ||
| Q. pyrenaica | Deciduous | GC-MS | Drought and CO2 | Aranda et al. [88] |
| Q. rubra | Deciduous | LC-MS | Drought | Top et al. [89] |
| HPLC-Orbitrap-MS | Drought | Suseela et al. [78] | ||
| Q. robur | Deciduous | TD-GC-MS | Operophtera brumata | Volf et al. [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escandón, M.; Castillejo, M.Á.; Jorrín-Novo, J.V.; Rey, M.-D. Molecular Research on Stress Responses in Quercus spp.: From Classical Biochemistry to Systems Biology through Omics Analysis. Forests 2021, 12, 364. https://doi.org/10.3390/f12030364
Escandón M, Castillejo MÁ, Jorrín-Novo JV, Rey M-D. Molecular Research on Stress Responses in Quercus spp.: From Classical Biochemistry to Systems Biology through Omics Analysis. Forests. 2021; 12(3):364. https://doi.org/10.3390/f12030364
Chicago/Turabian StyleEscandón, Mónica, María Ángeles Castillejo, Jesús V. Jorrín-Novo, and María-Dolores Rey. 2021. "Molecular Research on Stress Responses in Quercus spp.: From Classical Biochemistry to Systems Biology through Omics Analysis" Forests 12, no. 3: 364. https://doi.org/10.3390/f12030364